

3.发展差异研究中的“走出非洲”假说
3.1主要假说
关于发展差异深层原因的一类新兴研究提出了如下假说:当代各社会的祖先人群与东非人类摇篮之间的迁徙距离,影响了人际多样性在世界范围的分布,给发展造成了持续的驼峰状效应,反映了多样性对社会生产率水平有益与有害影响的权衡关系。(*本节的讨论部分参考了如下成果:Ashraf and Galor(2018)及Arbatlı et al.(2020)。)
该研究领域始自阿什拉夫和盖勒(Ashraf and Galor,2013a),他们指出人际多样性可能对总体生产率有方向相反的不同效应。多样性可以拓展个人特性的范围(包括技巧、能力与问题解决方法等),推动专业分工,激发创造性活动中的思想碰撞,加速对变化的技术环境的快速适应,从而促进经济发展。反过来,多样性也会扩大个人价值观、信仰、偏好与社会交往倾向的谱系范围,从而削弱相互信任与社会凝聚力,导致社会冲突和公共品供给效率下降,对经济发展不利。
所以,只要多样性与同质性对生产率的有利效应存在递减现象,人际多样性程度就理应会对经济发展造成驼峰状影响。具体来说,相比过于同质或者异质的社会,多样性水平适中的民族、国家或地区预计会有更高的经济发展水平。
3.2世界人口密度差异的起源
解剖学意义上的现代人类从非洲摇篮扩张的过程,给人际多样性水平在全世界不同群体中的差异留下了深远而不可磨灭的印迹。根据广为接受的“走出非洲”的人类起源假说,人类这一物种于大约30万年前在东非演化成解剖学意义上的现代形态,于距今约9万到7万年前开始分阶段迁徙历程,直至遍布全球各地。图2大致描述了人类走出非洲这一史前扩张过程的路线。
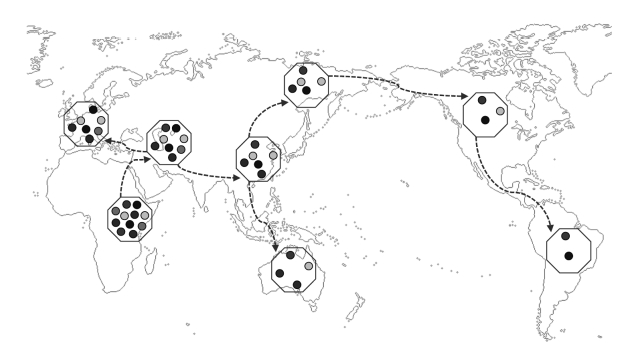 |
| 图2走出非洲的迁徙历程及其相关的顺序奠基者效应
注:本图描述了智人走出非洲的迁徙状况,及其对人际群体多样性分布的影响。在每次对外迁徙事件中,离开的(奠基者)群体都只带走了父辈殖民群体的一部分多样性,表现为奠基者群体的某个遗传特性的不同变异减少。图中的虚线代表大致的迁徙路径,小圆圈代表特定遗传特性的变异。 资料来源:Ashraf and Galor(2018)。 |
走出非洲的迁徙过程本质上决定了在更远离非洲的地区定居的群体,其多样性程度会更低。具体来说,存在一种与图2描述的情形类似的顺序奠基者效应:由于人类向世界其他地区的空间扩张是通过一系列独立步骤发生的,在每个步骤中,都有一个子群体离开父辈群体,去更远的地方建立新领地,由此只带走了父辈群体的一部分遗传特性。因此,如今在各个本地群体中观察到的多样性水平,均随着他们祖先与东非的迁徙距离增加而递减(参见Harpending and Rogers,2000;Ramachandran et al.,2005;Prugnolle、Manica and Balloux,2005;Ashraf and Galor,2013a)。
对应上述从东非起源的古代人群的瓶颈链条,图3的散点图描绘了迁徙距离对人与人之间基因多样性的影响,其中参考了人类群体遗传学研究文献涉及的全球207个代表性民族(Pemberton、DeGiorgio and Rosenberg,2013;Arbatlı et al.,2020)。(*这207个民族的样本包含Pemberton、DeGiorgio and Rosenberg(2013)的数据库的全部观测值,可以对应不同的民族祖居地,但不包括南美洲的苏芮人(Surui),他们被群体遗传学视为期望杂合度测量中极为特殊的例外(Ramachandran et al.,2005)。另外,即使把这一例外群体纳入样本,也不会改变结论的性质。)根据人类群体遗传学的研究,这些民族都属于目前所在地理位置上的本土居民,并在基因漂流上大致与其他民族分开。这些民族的空间分布参见图4。
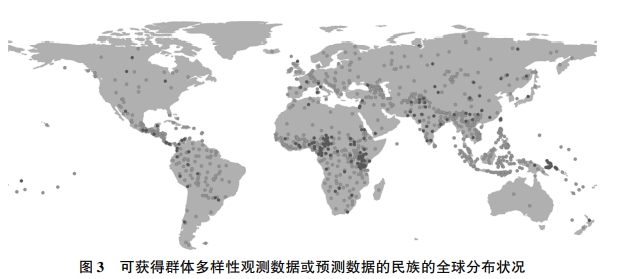 |
|
注:本图描述了群体多样性有观测数据或预测数据(根据迁徙距离测算)的民族在全球的分布状况。图中每个点代表一个民族的历史原生地的几何重心。深色点代表有观测数据的民族,浅色点代表有预测数据的民族。 资料来源:Arbatlı et al.(2020)。 |
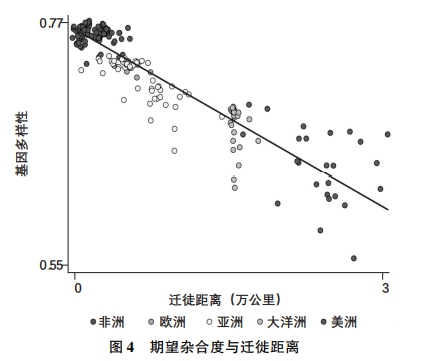 |
| 注:本图描述了迁徙距离对期望杂合度的负面影响,以207个民族样本为基础,数据来自Pemberton、DeGiorgio and Rosenberg(2013)。
资料来源:Arbatlı et al.(2020)。 |
为测算给定人群(如一个民族)中人们的基因构成多样性,群体遗传学采用了一种名为期望杂合度(expected heterozygosity)的指标,反映从人群中随机挑选出来的两个人在给定遗传特性范围内彼此不同的概率。具体做法是,首先根据给定遗传特性的不同等位基因或变异在人群中的占比,构建特定基因的期望杂合度指数,即随机选出的两个人在给定遗传特性上存在差异的概率。接下来通过测算大量基因或基因位点的杂合度,得出各基因位点的平均杂合度水平,即该人群的总体期望杂合度。因此,一个群体的期望杂合度Hexp的测算公式为:
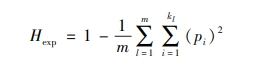 |
其中,m是考察的基因或基因位点的数量,kl是基因l自然产生的变异或等位基因的数量,pl是基因l的等位基因i在人群中的代表比例或出现频率。
群体遗传学在测算地理上的本土民族的期望杂合度时,采用了名为微卫星的特定类型基因点位的等位基因出现概率的数据。此类基因位点处于人类基因中的非蛋白质编组区域(即不会直接产生表型表达的区域),被视为具有选择中性的特点。在考察人际多样性如何影响不同人群的社会经济发展成就时,该测算指标具有一个关键优势:不会被在走出非洲之后作用于各人群的自然选择因素的任何未观察到的差异干扰,而这些潜在差异可能遮蔽祖先的顺序奠基者效应预测的关联关系。还有,差异选择及其背后的作用因素也可能影响社会经济发展成就,使我们难以识别人际多样性与社会经济发展之间的因果关系。
无论如何,要在理论上对社会经济发展成就产生影响,需要将中性遗传多样性的测算指标作为有效代理变量,用以反映表型表达特性和行为表达特性的多样性。事实上,来自体质人类学和认知人类学的大量证据显示,群体内的各种形态多样性与认知多样性在世界范围内确实存在起源于东非的古老顺序奠基者效应(Henn、Cavalli-Sforza and Feldman,2012),包括与颅骨特征(Manica et al.,2007;von Cramon-Taubadel and Lycett,2008;Betti et al.,2009)、牙齿特征(Hanihara,2008)、骨盆特征(Betti et al.,2013)、产道特征(Betti and Manica,2018)等有关骨骼特性的人际多样性,以及语言音素多样性(Atkinson,2011)等。(*还有,通过考察物质文化中包含的功能标记反映的群体内部多样性,学者发现了与人类在波利尼西亚群岛的最初扩张有关的顺序奠基者效应(Rogers、Feldman and Ehrlich,2009)。)下节的论述将指出,研究表明,采用祖先人群的迁徙距离作为预测群体多样性的代理变量,确实可以反映表型表达特性与行为表达特性的人际多样性对社会发展的潜在影响(Ashraf and Galor,2013a,2018)。关键在于,根据基因与文化共同演化及代际表观遗传的观点,群体多样性产生的社会经济影响有可能反映了“自然”与“培育”两方面特性的人际多样性的作用,以及这二者之间的相互作用。(*例如,有学者引入“遗传培育”的概念(Kong et al.,2018),指出与父母教育成就有关的父母基因型会影响子女的教育成就,即使后者并没有携带相关的等位基因变异,这表明原本来自“自然”的遗传特性可能通过“培育”机制实现代际传递。)
走出非洲的史前扩张历程除导致人类社会内部的人际多样性在世界各地呈现差异之外,还对社会之间的遗传分化程度产生了深远而持久的影响,后者在群体遗传学中采用名为FST遗传距离的指标测算。该指标反映的是:对任何两个人群,其综合遗传多样性中没有被相应的期望杂合度的群体加权平均值解释的部分。人群i和人群j之间的FST遗传距离由如下公式得出:
 |
随着各个群体在走出非洲的迁徙历程中逐渐分化,由FST遗传距离反映的群体之间的剩余遗传差异会因为如下因素而增加:(1)各个群体内部随着时间推移发生的随机变异,它会导致基因漂移;(2)不同的最终栖息地在环境决定的选择压力上存在差异。特别是,由于两个群体之间的迁徙距离部分反映了他们从共同祖先群体分化出来的时间长度,而且他们在后来相互建立联系的概率也随这一距离增加而减小,走出非洲假说的一个直接后果就是,两个群体的遗传距离随着他们之间的迁徙距离增加而扩大。图5利用选择中性的遗传标记(只反映随机变异导致的基因漂移的距离)测算的FST遗传距离,以散点图描述了各对群体之间的“距离隔离”导致的上述关系,其中包含53个民族的样本(研究数据来自HGDP-CEPH Human Genome Diversity Cell Line Panel;Cann et al.,2002;Ramachandran et al.,2005;Ashraf and Galor,2013a)。
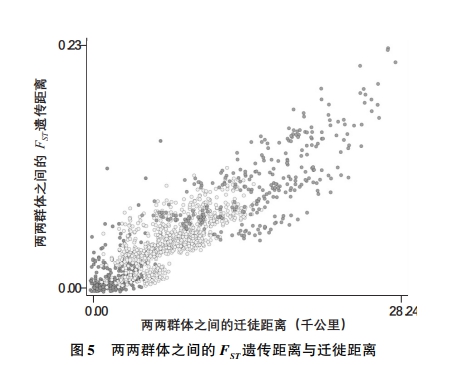 |
|
注:本图描述了两两群体之间的迁徙距离对遗传距离的正向影响,来自53个民族(HGDP-CEPH Human Genome Diversity Cell Line Panel)的1378个配对群体。黑色点代表两个民族归属世界同一地区(如都来自非洲、中东、欧洲、中亚和南亚、东亚、大洋洲、美洲)。浅灰色点代表两个民族归属世界不同地区,但都不包括大洋洲或者美洲。深灰色点代表两个民族归属世界不同地区,且至少一个来自大洋洲或者美洲。 资料来源:Ashraf and Galor(2013a,2018)。 |
3.3预测群体多样性
3.3.1国家层面的测算与识别
由于缺乏国家层面群体的人际多样性测算,民族层面的测算也很罕见,我们可以利用各个祖先人群的迁徙距离,及其同可观测遗传特性的人际多样性的强烈负相关,生成一种指数,以反映各个国家、地区或民族的群体多样性水平。具体来说,根据图3显示的联系,一个本土人群的人际多样性水平可以根据其祖先的迁徙距离预测。下文的讨论将为此提供更清晰的介绍。
测算国家层面的人际多样性的一个简单方法,是借助群体遗传学者的测算,把观察到的国内各个民族的期望杂合度指标简单加总。然而,这一方法存在某些重大缺陷。第一,虽然有已观测群体多样性(observed population diversity)的民族具有全球代表性,在空间上遍布世界各个大陆和地区,却只跨越一部分国家。因此,这一简单方法只能对有限的国家样本得出已观测群体多样性。第二,对一国人口中各个民族的期望杂合度做简单(或人口加权)平均,并不能反映各民族两两之间的生物文化距离带来的其他多样性。此问题在新大陆尤其突出,那里各国人口中包含的各民族,其祖先来自地理上相距遥远的各个地区。第三,已观测群体多样性与民族层面的社会经济发展成就的潜在内生联系意味着利用这一简单测算方法可能使国家层面的多样性测算受到反向因果关系或者被遗漏变量的干扰。
为克服样本局限、对中性遗传标记的依赖,以及在简单方法下采用可观测群体多样性的内生影响,阿什拉夫和盖勒(2013a)开创的研究领域采用了如下策略,以利用迁徙距离对世界各民族的期望杂合度差异的强大解释力(类似于图3描述的情形)。具体来说,第一步是利用HGDP-CEPH样本中的53个民族的期望杂合度对迁徙距离的回归系数,针对世界各地的所有前殖民社会,根据它们在公元1500年时所处的地理位置,得出一个已预测人际多样性指标。利用更新和更大的数据库(来自Pemberton、DeGiorgio and Rosenberg,2013),而非HGDP-CEPH样本,也能得出基本一致的结果。这里的关键是,在发现新大陆与殖民时代重大洲际移民行动之前,历史上的各个社会所处的地理位置基本上反映了他们的祖先人群在史前走出非洲的迁徙行动结束时最终抵达的地方,所以,前殖民社会的多样性应该主要取决于源自东非的顺序奠基者效应。下一节还将阐述,与远古时期走出非洲的迁徙路线有关的距离外生性意味着,已预测群体多样性(predicted population diversity)的这一指标更适合用来识别人际多样性对前殖民时代各社会经济与制度相对发展水平的影响。
但无论如何,为识别多样性对现代世界各国经济发展与社会政治制度差异的影响,我们有必要为已预测群体多样性构建一个指标,包含现代各国人口中多个民族祖先带来的其他多样性,反映过去500年左右的大规模跨洲与跨地区移民。因此,阿什拉夫和盖勒(2013a)设计了现代国家人口的多样性预测指标,其测算基础包括:(1)各民族在一国人口中的代表比例;(2)每个民族自身的群体多样性,由其祖先的迁徙距离预测得出这些次国家群体(subnational group)在人口中所占比例的数据来自Putterman and Weil(2010)的“1500—2000年世界迁徙矩阵”(World Migration Matrix,1500-2000),该数据库包含每个国家在公元2000年的人口中,祖先来自公元1500年的其他国家的各个人群所占的比例。;(3)这些民族之间的两两生物文化距离,由各自祖先人群之间的迁徙距离预测得出。
具体而言,根据之前对FST遗传距离的定义,设想某个现代国家的人口包含两个民族,其祖先来自A和B两个不同地理位置,则该国人口的总体多样性水平可计算如下:
 |
元1500年之后的殖民时代从地点i迁移来的民族在该国人口中的占比。此外,
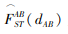 |
是地点A和B之间的迁徙距离预测的生物文化距离,利用类似于图5的回归线的对应相关系数计算得出。实际上,由于现代国家的人口构成通常远不止两个民族,上述步骤需要针对国家人口中的大量组成民族做反复运算。
现代国家人口的群体多样性测算既包含民族内多样性,又包含民族间距离,使它在识别群体多样性对各国现代经济发展与社会政治制度差异的作用上非常有价值。相比之下,试图反映各国人口中民族分化(ethnic fragmentation)程度差异的其他常用多样性测算指标,通常没有利用各国人口中不同民族语言群体所占比例之外的信息,也就是说,它们隐含地假设这些民族具有内部同质性,并且相互之间的生物文化距离都相等。(*对民族分化的更精细的测算指标,例如(1)反映“文化多样性”的Greenberg Index(来自Fearon,2003;Desmet、Weber and Ortuño-Ortín,2009);(2)民族语言极化指数(来自Desmet、Weber and Ortuño-Ortín,2009;Esteban、Mayoral and Ray,2012),该指数包含了两两之间的语言距离,而这一语言亲近度在层级语系树中随任何两种语言之间的共同分支数量而单调增加。然而,此类信息受制于层级语系树的性质,位于相同分支层级的语言必然有着相同的距离。)而利用合理外生迁徙距离(各国祖先人群与东非之间及相互之间的迁徙距离)来预测民族内部的多样性和民族间的距离,除考虑了民族内的多样性及相互距离外,还可以帮助识别各国人口多样性对当代发展成就的影响,甚至适用于各组成民族的已观测期望杂合度与FST遗传距离没有现成数据的国家。
还有,由于公元1500年以来的跨地区移民以及各国人口中的民族比例可能受发展和长期空间格局冲突的影响,现代各国人口的多样性对其社会发展成就或许具有内生关系。为解决此问题,该研究领域的某些论文采用了如下两种实证策略。第一种策略是把实证分析局限于归属旧大陆(非洲、欧洲和亚洲)的国家样本,这些国家的人口多样性主要反映了远在殖民时代之前就生活在那里的土著人群的特征。这一策略基于如下现象:旧大陆自公元1500年后的人口流动并未导致过去彼此相距遥远的群体显著混合。第二种策略则是利用一个两阶段估计量来考察具有全球代表性的国家样本,把某个国家的史前土著人群的迁徙距离作为其现代国家人口多样性的工具变量。该策略的基础是识别如下假设:一个国家的史前土著人群的迁徙距离对该国当代全体人口的经济和社会政治发展成就可能是外生因素。
3.3.2民族层面的测算与识别
关于群体多样性对经济发展与社会政治制度差异的作用的研究项目,在考察群体多样性与民族层面的经济、政治与社会成就的联系时,采用了多种策略来减轻反向因果关系、被忽略的文化地理与人类特征以及分类选择偏向的干扰。在人类发展历史上,不同民族的经济发展成就和内部冲突频率的差异可能改变了这些民族内部观测到的人际多样性。因此,某个民族已观测的群体多样性与群体生产率或内部冲突频率的联系,可能部分反映了反向因果关系。此外,民族层面的这些联系可能部分受到被忽略的文化、地理或人类特征的影响。为缓解这些潜在疑虑,该研究领域的实证分析利用当代本土民族的已观测群体多样性与迁徙距离之间的强烈负相关——这源自之前强调的古老顺序奠基者效应(Harpending and Rogers,2000;Ramachandran et al.,2005;Prugnolle、Manica and Balloux,2005;Ashraf and Galor,2013a)——为具有全球代表性的900多个民族样本的已预测群体多样性构建了一个指标。
当然,仍有几种场景可能从理论上削弱这一方法的可信度。首先,走出非洲的选择性迁徙或者在古代迁徙路径上发生的自然选择都可能影响人类特性,造成民族层面的发展成就与迁徙距离对人类特性之多样性的影响无关。迁徙距离虽然与人类遗传特性的多样性显著负相关,但与一个群体特性的平均水平似乎无关,例如身高、体重、皮肤反射水平等,后者更多取决于和赤道的距离(Ashraf and Galor,2013a)。其次,迁徙距离可能与重要历史位置(如技术前沿所在地)的距离有关,因此可能通过发展过程与冲突事件中的其他距离产生影响,而不是通过影响群体多样性来发挥作用。不过在考虑了迁徙距离之后,研究发现与公元1年、1000年和1500年的技术前沿地区的距离并没有从根本上改变已观测群体多样性对群体生产率或内部冲突水平的影响(Ashraf and Galor,2013a;Arbatlı et al.,2020),从而进一步证实,可以利用与走出非洲的迁徙历程有关的顺序奠基者效应,来识别群体多样性对民族层面各种发展成就产生的影响。
还有,如果走出非洲的实际迁徙路径与地理特征(例如土地质量、崎岖程度、气候条件、贸易条件等)有关,那么这些特征可能直接影响创新活动与内部冲突,从而给识别带来麻烦。当然,这要求有利于发展成就的地理因素与走出非洲的主要迁徙路径保持一致,只要每个主要迁徙分叉都源自这条主要迁徙路径。具体而言,这要求在迁徙路径的几个关键分叉处(例如肥沃新月地带以及与之相关的向东迁往亚洲和向西迁往欧洲),有利于发展的地理因素的影响必须沿着分开的次级迁徙路径对称地递减,而这在现实中不太可能出现。总之,本研究领域的分析表明,在考虑到民族祖居地、空间依赖等若干带来干扰的潜在地理特征,以及未观察到的非时变地区间差异之后,已预测群体多样性对民族发展成就的影响的性质未受影响,由此识别出了世界同一地区不同民族的人际多样性与发展成就之间的相关性。
我们观察到的群体多样性与民族发展成就的相关性,还有可能是因为把不够多样化的群体划分为地理上的微小单元,他们爆发冲突的风险较低,但也不太有利于促进创新活动。这种划分虽然不会否认群体多样性同冲突或创新活动之间存在正相关,却会削弱对这些相关性的理论解释。不过,这种划分要求事前的冲突风险与创新潜力的空间分布必须与迁徙距离负相关,导致冲突或创新的地理特征也必须与走出非洲的主要迁徙路径、每个主要分叉以及次级路径都负相关。这些要求在本质上极不可能实现,但学者们仍然考虑了各民族祖居地的一系列地理特征的异质性、空间的自相关关系以及世界各地的地区固定效应,从而进一步减轻划分更小单位造成的干扰问题。
3.4实证发现
3.4.1各国的群体多样性与相对发展成就
阿什拉夫和盖勒(2103a)借助国家层面的人际多样性测算数据,实证分析了多样性水平对社会生产率的有益影响与有害影响。他们的发现与假说一致,即主要由史前走出非洲的迁徙历程的顺序奠基者效应决定的人际多样性,确实对人均收入水平产生了显著的驼峰状影响,可以解释2000年全世界各国生活水平差异的16%。
虽然该研究的焦点是当代的相对发展水平,但也证实多样性对经济发展的驼峰状影响在历史上和当代都存在,表明各社群内部的多样性远在工业革命到来前就决定了相对发展状况。在前工业化时代,相对发展水平由马尔萨斯机制主宰,其含义是,社会生产率进步主要表现为推动人口的增长,而非人均收入的提高。在这个时代,较发达社会的特征是更高的人口密度,而不是更高的生活水平(Ashraf and Galor,2011)。所以,上述研究的历史分析着眼于解释各个社会在公元1500年时的人口密度差异。
该研究利用对殖民前社会的预测多样性,以及遍布全球的观测样本,揭示了人际多样性对公元1500年时的生产率(表现为人口密度)产生的驼峰状影响。特别是,这里考虑了潜在混淆因素的干扰,例如各社会发生新石器革命的时机有差异,与历史发展有关的各种地理因素,以及未观察到的不同大陆的非时变差异等。该发现对大量敏感性检验保持了稳健性,“安慰剂检验”也显示:采用与东非之间的航线距离或者与其他地理位置的迁徙距离时,就不会出现类似的驼峰状影响。
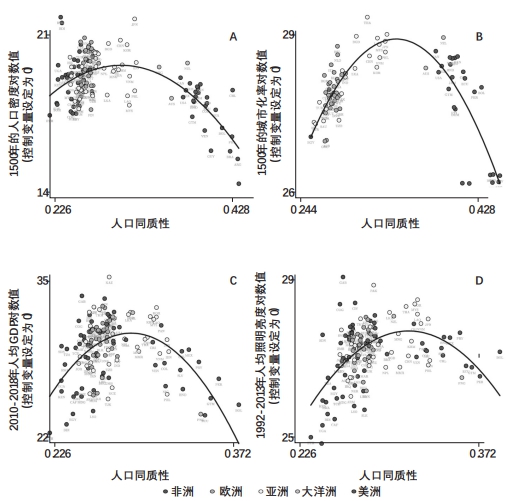
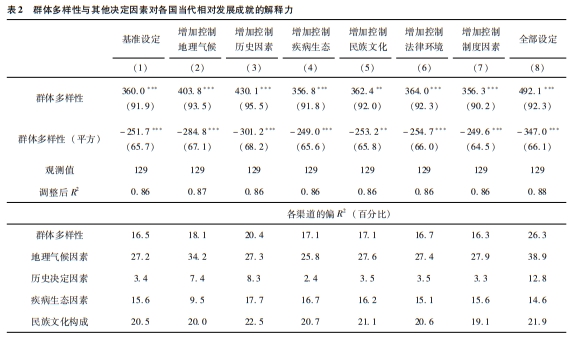
表1的第1列和第2列利用更新的矢量数据信息重现了上述结果,其中考虑了地理、气候和历史制度的特征,新石器革命与最早人类定居发生的时间,以及世界各区域的固定效应等。第2列考察的人口密度的所有模型设定也揭示出明显的驼峰状关系,相比最低或最高的多样性水平,预测拐点附近的多样性水平对应的人口密度要高出3.3~0.9个对数点。这一关系见图6A。另外,表1的第3列和第4列利用类似的条件设定,表明如果把公元1500年时的城市化率作为前工业化社会相对发展的代理指标,人际多样性对生产率水平的驼峰状影响依然存在。图6B显示了第4列估计的驼峰状回归关系。
 |
阿什拉夫和盖勒(2013a)利用现代各国群体多样性测算指标,发现多样性对公元2000年时的人均收入有显著的驼峰状影响。该发现考虑了新石器革命在各国发生的时机差异,与现代经济发展有关的各种地理、文化与制度关联因素,以及各大洲未观察到的非时变特征。这一关系在控制了公元1500年的人口密度后依然稳健,说明多样性的驼峰状影响并不仅仅反映经济发展的长期惯性。另外,如果把样本限于自前殖民时代以来主要人口仍为本地土著群体的国家,该关系依然成立,因此可以减轻过去500年以来地区间人口流动的内生性问题。
人际多样性对当代各国的相对发展水平有显著的驼峰状影响,这在表1的第5列和第6列再次出现,这里采用的全部设定(full specification)是在之前各列中加入当代面临的疟疾风险,以及与当今发展成就有关的大量文化和制度协变量,并把2010—2018年的人均收入水平作为因变量。图7C显示,第6列考察的人均收入的全部设定得出了如下预测:相比多样性水平的最低点和最高点,拐点附近的多样性水平对应的人均收入分布高出2.1和1.2个对数点。
 |
|
图6群体多样性与各国的相对发展成就 注:本图描述的是群体多样性对各国当代经济发展成就的驼峰状影响,其中控制了地理、气候、文化和制度特征,新石器革命与最初人类定居出现的时间,以及世界各区域的固定效应。上两图描述的是预测群体同质性(即1减去由迁徙距离预测的群体多样性)对公元1500年时经济发展的影响,分别为人口密度对数值(A)与城市化率对数值(B)。下两图描述的是预测祖先调整后的同质性(即1减去由一国各祖先人群的迁徙距离以及他们之间两两迁徙距离预测的群体多样性)对当代经济发展成就的影响,分别为2010—2018年人均收入对数值(C)与1992—2013年人均照明亮度对数值(D)。每个图都展示了二次拟合得到的增加部分的残差,对应表1中的相应偶数列。 |
考虑到现代世界的人均收入水平测算可能受到干扰,尤其是在欠发达经济体,有学者指出,如果用调整后人均夜间照明亮度(1992—2013年平均值,可以通过卫星太空监测得出)来反映各国的相对繁荣水平,多样性对发展成果的驼峰状影响依然成立(Ashraf、Galor and Klemp,2014)。表1第7列和第8列介绍了这一分析结果,对之前两列考察人均收入的设定做了更新。图7D描绘了第8列的全部设定估计得出的显著驼峰状关系。这些发现可以减轻如下问题,即利用人均收入测算生产率时,高多样性对总生产率的不利影响可能受到向上扭曲,因为鉴于多样性对社会凝聚力有不利影响,在多样性较高的社会里,影子经济的规模可能会大得多。
群体多样性的相对重要性与其他基本决定因素
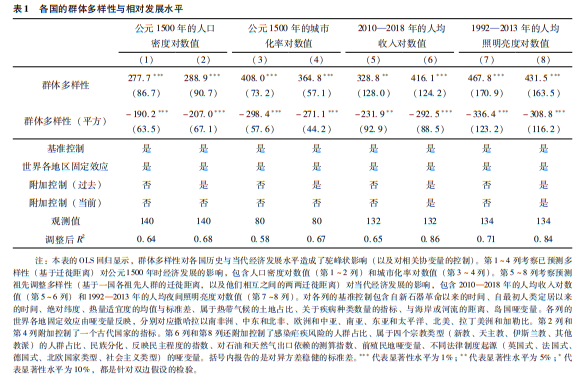
本节将估计人际群体多样性与其他6个基本因素,即地理因素、政治制度、民族文化构成、历史因素、疾病生态、法律环境,对当代各国经济繁荣水平(用2010—2018年平均人均收入表示)的相对贡献。具体来说如表2所示,该分析考察了与这些因素对应的各组协变量的附加解释力(偏R2统计量)。
分析过程首先是第1列的基准设定情形,6个基本因素分别由一组核心协变量代表。但为确保对所有因素做平衡的评估,相比基准设定情形,随后的设定(即第2列至第7列)采取了一次扩展一组协变量的办法。最后考察全部设定的情形,其中所有影响因素都由之前各列中扩展的各组协变量表示。(*本分析考察的各个因素在基准设定情形和各个扩展设定情形中使用的变量列表,见表2的注释部分。)
从结果看,与人际群体多样性的驼峰状影响有关的偏R2统计量表明,对于各种模型设定,这一因素可以解释当代人均收入剩余差异的16%~26%(控制其他基本因素之后)。在其他基本因素中,地理气候因素的偏R2为26%~39%,反映法律环境、民族文化构成、疾病生态等因素的偏R2分别为13%~27%、19%~23%和10%~18%。另一方面,历史决定因素(从新石器革命以来的时间长度、自人类最初定居以来的时间长度、古代国家状况与殖民历史等)以及可以尽量减小剥夺风险的政治制度因素(对行政权力的限制与民主化程度等)的偏R2分别为2%~13%和2%~8%。最后在包含全部设定的第8列纳入了所有因素的协变量组,纳入人口密度可以增加26%的解释力,地理气候因素可增加39%,民族文化构成为22%,疾病生态为15%,法律环境与历史决定因素各为13%,政治制度因素为8%。
 |
 |
3.4.2不同民族祖居地的群体多样性与相对发展成就
阿什拉夫、盖勒和克莱姆普(Ashraf、Galor and Klemp,2020a)实证分析了群体多样性对民族层面的生产率的影响,同时考虑了多种因素可能产生的混杂影响(confounding effects),这些因素包括观察到的各民族特有的地理、文化和制度差异,以及未观察到的非时变区域特征差异等。该研究发现,世界范围的207个民族的观测多样性(Pemberton、DeGiorgio and Rosenberg,2013)以及全球900多个民族的预测多样性(基于迁徙距离),对肥沃新月地带开启新石器革命以来的整个人类发展历程中的经济繁荣水平都有显著的驼峰状影响。
该结果表明,不同民族祖居地(ethnic homeland)的人际多样性差异导致了他们的极长期经济发展差异,例如历史上不同时点(公元前10000年到公元1500年之间每个千年的起始年份)的人口密度,以及当代调整后的人均夜间照明亮度。
表3介绍了阿什拉夫、盖勒和克莱姆普(2020a)的部分发现。利用不同民族祖居地的差异,他们的回归分析显示,人际多样性对公元前5000年(第1~2列)、公元前3000年(第3~4列)、公元前1000年(第5~6列)、公元100年(第7~8列)的人口密度有显著的驼峰状影响,对1992—2013年调整后的人均夜间照明亮度也有类似影响(第9~10列),其中利用了观测多样性(奇数列)与预测多样性(偶数列)。分析历史人口密度的回归考虑了人类最早定居的时间以及大量地理和气候因素,加上世界各地区的固定效应;而分析当代夜间照明亮度的回归则控制了如今的疟疾风险以及当代文化和制度协变量的影响。
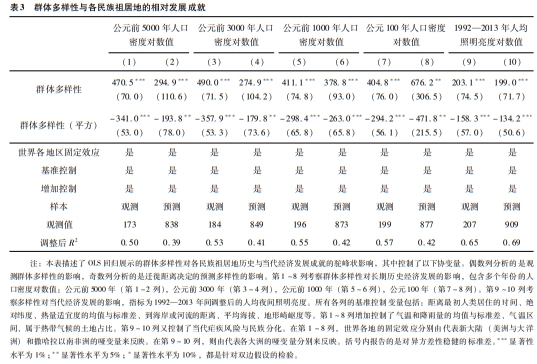
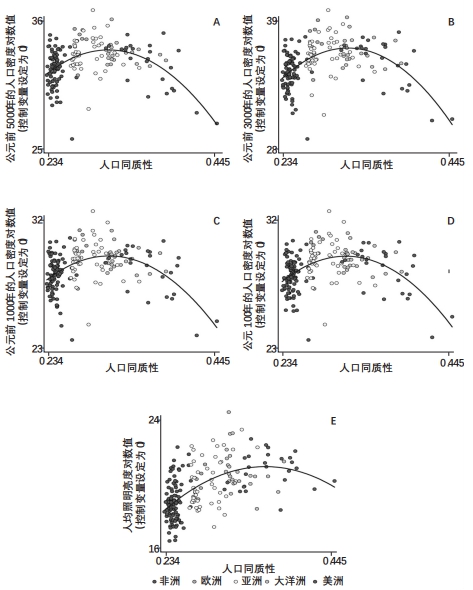
为解释表3揭示的显著驼峰状影响,可以看第5列的回归对人口密度得出的预测:相比多样性程度的最低点和最高点,拐点附近的多样性水平对应的历史人口密度分别高出5.4和1.9个对数点。此外,第9列的回归预测:相比多样性程度的最低点和最高点,拐点附近的多样性水平对应的当代夜间照明亮度分别高出1.2和2.5个对数点。表3的奇数列回归揭示了观测多样性对不同民族祖居地在不同时期的相对发展水平有驼峰状影响,这些影响按顺序展示在图8中。
阿什拉夫、盖勒和克莱姆普(2020a)对民族层面的分析提供了最优场景,借助以往研究记录的200多个民族的观测多样性数据(Pemberton、DeGiorgio and Rosenberg,2013),证实了阿什拉夫和盖勒假说的主要预测的稳健性。
 |
 |
| 图7群体多样性与民族祖居地的相对经济发展成就
注:本图描述了人际群体多样性对各民族祖居地历史与当代经济发展成就的驼峰状影响,其中控制了地理、气候、文化与制度特征,距离最早人类定居的时间,以及世界各地区的固定效应。前四幅图描述了观测群体同质性(即,1减去观测群体多样性)对长期历史经济发展的影响,指标分别为各时间点的人口密度对数值:公元前5000年(A)、公元前3000年(B)、公元前1000年(C)、公元100年(D)。最后一幅图描述了群体同质性对当代经济发展的影响,指标为1992—2013年调整后的人均照明亮度对数值。每个图都展示了二次拟合得到的增加部分的残差,对应表2中的相应偶数列。 |
后者的分析只采用了HGDP-CEPH的53个民族样本。此外,人际多样性对前殖民时代与现代的相对发展水平的影响,在国家层面的次优情形中也是稳健的。实际上,借鉴阿什拉夫和盖勒(2013a)的方法,我们可以基于迁徙距离对具有全球代表性的国家样本测算出预测群体多样性。从HGDP-CEPH样本与后来的扩展样本中得出的迁徙距离与观测多样性之间关系的估计系数几乎相同,这说明预测多样性对各国历史与当代发展成就差异的驼峰状效应不受样本选择的影响。(*第三优的方法是对国家层面的观测多样性与经济发展水平之间的关系做非推理性的初步考察。采用该方法的一项研究提出(Rosenberg and Kang,2015),观测多样性与历史人口密度的驼峰状形态在39个样本国家中没有统计显著性,其依据是各国内部观测到的某些民族子集的期望杂合度的平均值。然而,利用此方法检验人际多样性对经济发展水平的驼峰状影响存在根本缺陷,原因有二:其一,由于一个国家的观测民族子集未必对全国具有代表性,且样本中包含的民族只覆盖了39个国家(没有全球代表性),该研究在国家层面存在严重的样本选择偏差。也就是说,基于不具代表性的国家样本,观测多样性与经济发展水平在各国之间的驼峰状联系是否存在的假设无法被证实或拒绝。其二,观测多样性或许反映了过去的社会经济成就,例如地区内部的社会冲突和移民状况,它们本身受过去经济发展状况的影响。于是,该研究将受到反向因果关系与遗漏变量的困扰,使多样性与经济繁荣之间的驼峰状关系被遮蔽。的确,Ashraf、Galor and Klemp(2020b)的研究利用不具代表性的国家样本发现,一旦控制住足够多的潜在地理混淆因素的作用,观测多样性与历史人口密度之间就会出现显著的驼峰状关系。Ashraf、Galor and Klemp(2020a)指出,如果采用有效的统计方法分析扩展后的民族样本遗传数据,阿什拉夫和盖勒假说的主要预测就非常稳健。)
3.4.3群体多样性发挥影响的机制
人际多样性对生产率产生驼峰状影响的简化理论认为,多样性可能通过几种机制影响经济发展成就,反映了多样性的成本与收益权衡的不同因素。本节将回顾这些机制的相关证据,它们来自该研究领域的各类研究文献。
反映多样性对社会凝聚力产生消极影响的机制
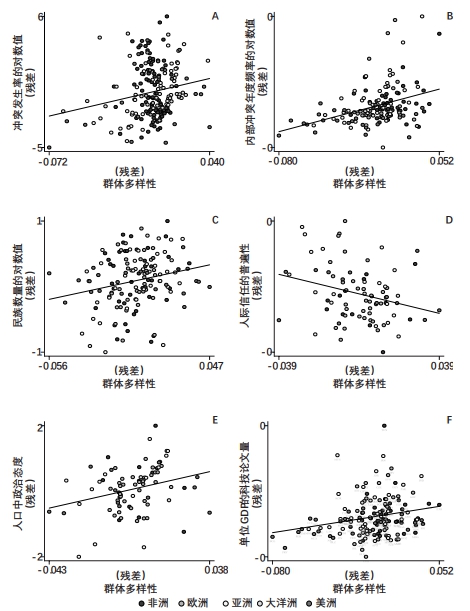
对内部冲突的影响。社群内部的冲突反映了一种基本机制,某个群体的人际多样性可能通过它迟滞经济发展。(*关于冲突与经济发展落后之间多层面联系的文献综述,可参见Ray and Esteban(2017)。)有学者考察了各国与各民族的差异,发现主要由走出东非的古老迁徙历程决定的群体多样性,是历史和当代内部冲突风险与剧烈程度的关键影响因素(Arbatlı et al.,2020),这里控制了与冲突相关的各种地理、文化和制度因素,经济发展成果,以及各大洲或各地区独有的未观察到的非时变特征。重要的是,与经常采用的民族分化和极化指标不同,人际多样性反映了群体内部与群体之间在个人遗传特性上的差异,它不仅能解释群体之间的冲突,还适用于群体内部的冲突。该研究进一步发现,人际多样性可能通过其他渠道影响一个社会的内部冲突,包括国家层面的民族分化程度提高、人际信任度降低,以及对公共品和再分配政策的偏好出现更严重分歧等。根据该研究的结果,图8描绘了:(1)观测群体多样性对1989—2008年各民族祖居地内部冲突发生率的正向影响(图8A);(2)当代各国的群体多样性对1960—2017年各国内部冲突年度发生频率的正向影响(图8B)。
对民族文化分化的影响。之前的若干研究文献认为(Easterly and Levine,1997;Alesina et al.,2003;Alesina and La Ferrara,2005),国家层面的民族多样性程度(以各民族分化与极化程度测算)同各种维度的经济发展落后相关。有项研究考察了这一因果链条的深层来源,发现史前决定的人际多样性可能是各国人口中民族语言分化的根本原因之一(Ashraf and Galor,2013b)。具体来说,该假说认为在远古走出非洲的迁徙历程之后,给定地点的初始群体多样性禀赋(endowment)可能通过一个内生的民族选择过程促进了当地不同民族的形成,反映了每个民族的规模与内部凝聚力之间的权衡关系。规模较大的民族可以受益于规模经济,却更难开展协调行动,因此可能更缺乏凝聚力。再考虑到人际多样性增加可能削弱民族内部的凝聚力,给定地点的初始群体多样性禀赋较大可能导致更多数量的民族出现。假以时日,“文化漂流”和文化标志的“偏向传播”(例如方言、习惯、传统、社会行为规范等)会把民族中的“内部人”与“外部人”区分开来,民族之间在这些文化标志上的分化就会愈加突出,最终导致不同的集体身份认同沿着民族界限被确立起来。
与该假说一致,研究发现国家层面的人际多样性对各种民族语言多样性指标具有强烈的正向影响,这里考虑了诸多因素的潜在混杂影响,如新石器革命发生的时机、距离人类初始定居的时间长度、殖民历史、民族多样性的地理决定因素、未观察到的各大洲独有的非时变特征等。此外,为解决因果关系的识别问题,对于只包含旧大陆国家的样本,上述发现依然成立,基本上不受殖民时代洲际移民的内生性的影响。同样,对于全球样本国家,如果采用前殖民时代一国土著人群的史前迁徙距离作为当代群体多样性差异的可能外生来源,上述发现也具有稳健性。人际多样性对国家层面民族数量的正向影响(Ashraf and Galor,2013b;Arbatlı et al.,2020),在图8C给出了直观的描述。
 |
|
图8群体多样性对总生产率的各种影响机制 注:本图描述了各民族祖居地和各国人际群体多样性通过(多样性影响总生产率的)不同机制产生的影响。A描述了各民族祖居地的观测群体多样性对1989—2008年内部冲突普遍性的影响,其余部分描述了祖先调整后的预测多样性对各国的影响:(1)各国在1960—2017年发生内部冲突的年度频率(B);(2)民族的数量(C);(3)世界价值观调查中反映的1981—2008年人际信任的普遍性(D);(4)世界价值观调查中反映的1981—2008年的个人政治态度(由政治上的左右评分表示)的国内分布(E);(5)2010—2018年单位实际GDP对应的科技论文的数量(F)。每幅图都展示一个增加变量的图示,以反映考虑到与解释结果有关的控制变量矢量之后的关系。 资料来源:Ashraf and Galor(2013a,2013b,2018);Arbatlı et al.(2020)。 |
对人际信任度的影响。人际多样性可能导致个人价值观、信仰、偏好与倾向性的频谱范围扩大,从而削弱社会资本,拖累经济发展。与该假说相符,研究发现当代的群体多样性对一般人际信任在各国的普遍程度有显著的负面影响(Ashraf and Galor,2013a;Arbatlı et al.,2020),其数据来自1981—2008年的世界价值观调查。(*有关实证研究综述参见Algan and Cahuc(2014),这些研究证实了一般信任程度与经济成就的因果关系,并考察了数据中展现的各种影响机制。)如图8D所示,这一关系考虑了各国的异质性(与经济发展和民族多样性有关的各种地理协变量)以及未观察到的世界各地的固定效应。
另有研究利用考察个人间差异的两个独立分析,发现个人之间信任缺失的普遍性与祖先人群的人际多样性水平显著相关(Arbatlı et al.,2020)。
•第一个分析考察非洲晴雨表调查(Afrobarometer surveys)得到的个人间不信任水平同他们非洲祖居地的观测群体多样性的联系,无论这些受调查者目前是居住在祖居地,或是已迁徙到不同的地方。在考虑如下多种潜在混杂因素的矢量后,这一相关性依然具有稳健性:(1)目前居住国的固定效应;(2)个人特征(年龄、性别、教育、职业、生活环境和宗教等);(3)祖居地受到奴隶贩运影响的程度;(4)居住地特征(学校、电力、自来水、排水系统、诊所等机构的存在,城市居民身份等);(5)祖籍国的固定效应;(6)目前居住地与祖居地当前的民族语言多样性水平。
•第二个分析考察一般社会调查(General Social Survey)收集的美国第二代移民的不信任水平,同其父母原籍国当代群体多样性的联系。聚焦一国的移民,可以很自然地控制移民接收国各种未观察到的非时变特征(包含地理、文化和制度等方面)。此外,该研究还明确考虑了多种个人层面的混淆因素,以及地理特征、世界各地的固定效应、民族语言分化和极化程度的影响(都是对应他们的父母原籍国)。对政治偏好差异的影响。个人在价值观、信仰和一般偏好方面的差异尤其可能导致对公共品类型的偏好差异,以及个人对经济不平等和政府再分配政策的倾向发生分歧,从而引发政治极化与制度扭曲,带来社会冲突和经济落后。对于这一机制,有关研究指出(Ashraf and Galor,2018;Arbatlı et al.,2020),当代的群体多样性对各国的政治偏好异质性产生了显著正向影响,表现为各国内部由个人报告的政治“左右”天平上所处位置的差异,其数据来自1981—2008年的世界价值观调查。图8E描绘了这一联系,同时控制了与发展和政治经济有关的各种地理协变量,以及未观察到的世界各地的固定效应。
对攫取性制度的影响。专制社会治理模式的出现与延续,是人际多样性可能导致当代经济发展落后的另一个关键机制。有学者提出的假说认为,史前确立的人际多样性可能提高社会对某些正式制度的需求,以抑制多样性对社会凝聚力的不利影响,但同时,多样性对社会内部的经济不平等与阶层分化的作用可能使制度发展走向更具攫取性的专制治理模式(Galor and Klemp,2017)。该研究借助前殖民地时代各民族祖居地的差异,发现在控制若干地理因素与各大洲未观察到的非时变特征等混杂效应之后,人际多样性仍对前殖民时代专制制度的普遍程度具有正向影响。这可能反映了多样性的双重作用:对正式制度的需求以及社会阶层分化的出现。此外,该研究还指出人际多样性在全世界的空间差异可能导致专制制度在当代各国之间的不同,这部分源于制度、文化和人口结构特征的长期延续。
反映多样性对生产率产生有利影响的机制
对创新活动的影响。人际多样性可以拓宽相互补充的不同认知方法,以解决问题和生产知识,从而促进创新活动。针对这一假说,有研究指出当代的群体多样性与各国在1981—2000年的年均科研论文发表数量存在显著正向关联(Ashraf and Galor,2013a)。该发现考虑了潜在混杂因素的影响,包括各国发生新石器革命的时机差异,与当代经济发展有关的各种地理、文化和制度协变量,以及各大洲未观察到的非时变特征等。图8F描述的是,当代各国的群体多样性对2010—2018年单位实际GDP对应的科研论文数量的类似正向影响,其中控制了地理和气候因素,以及各地区未观察到的非时变特征等。(*人际多样性对经济发展的有益作用也得到了其他研究的实证支持,如Ager and Brueckner(2018)。该研究考察了美国各州在19世纪后期的差异,发现由于欧洲移民的到来、多样性得到更大提升的县级群体,在1870—1920年获得了更高的收入增长率与人均科技专利增长率。另一项研究(Delis et al.,2017)借助北美与英国多家股票市场上市企业的面板数据,发现在董事会中加入一些原籍国有不同遗传多样性水平的董事,会提升企业业绩。该研究推测,其发现反映了对生产率有利的文化、心理、生理及其他遗传特性的人际差异,这些特性差异尚未反映在其他多样性测算指标中。)
对劳动分工与贸易收益的影响。近期的一篇论文(Depetris-Chauvin and Özak,2020)实证考察了如下假说(Ashraf and Galor,2013a):人际多样性可以通过扩大个人的技术、能力和认知方法的频谱范围,促进社会的劳动分工。该研究利用前殖民时代不同民族祖居地的观测差异,发现与上述假说相符,史前确立的人际多样性对社会中不同生产活动的专业化程度可能有正向影响,从而强化了参与市场交换并从中获得经济利益的倾向。(*我们可以想到,在前工业化生产中,物质环境与个人能力形成了互补的投入品。有趣的是,该研究还发现,农业生态与农业气候的异质性程度强化了人际多样性对前殖民地社会出现的经济专业化的正向影响。)该研究还指出,前殖民时代经济专业化程度更高的民族祖居地往往在当时和现在都有显著更高的经济发展水平,这部分是源于更为丰富的人际多样性。
通过思想交流对认知发展和人力资本培育的影响。有意思的是,在比国家或民族低得多的层面上,同样发现了人际多样性对生产率的积极影响。有学者考察了美国威斯康星州具有民族同质性的各所高中的差异,发现1957年高中学生群体的期望杂合度可能对毕业生后来的经济成就(包括完成的学业年限、第一份工作的职业声望、1974—1992年的家庭收入水平等指标)具有显著的正向影响(Wisconsin、Cook and Fletcher,2018)。该研究还提供了与一种新的行为机制相符的证据:身处更为多样性的学校教育环境,可能让一个人的性格更容易接受创造性,对新奇思想更开放,从而使他在人生中取得更大的社会经济成就。关键在于,由于该研究采用的数据包含的高中生群体完全是由欧洲祖先的后裔构成,这些群体内部的文化同质性与社会凝聚力使得该分析主要反映了多样性的好处,而不是代价。另外,由于这些发现是基于对州内差异的分析,理应不会受到各国之间(甚至国家内部的各州之间)混杂因素的干扰。
3.5政策启示
若干研究发现人际多样性(由预测的群体多样性表示)对经济发展成就的不同影响存在基本权衡关系(Ashraf and Galor,2013a;Ashraf、Galor and Klemp,2020a)。根据迁徙距离得出的预测多样性水平是经济发展的深层决定因素,但这并不表示一个群体的遗传特性完全主宰了其经济命运。迁徙距离通过与地理、生物和文化特征的相互作用影响人际多样性,因此政策也可以着眼于调整这些作用渠道。
具体而言,多样性对生产率的影响意味着,一个社会可以采纳恰当的政策,以发挥其人口的现有群体多样性的积极影响,缓解其消极影响,从而改变群体多样性影响社会经济发展的环境。过度多样性的社会可以重点鼓励公民的社会参与,提高政治制度的品质,改变公共品提供的低效和扭曲,以培养人际信任,缓和潜在冲突。过度同质性的社会则可以着眼于促进技能、职业和培训项目的多元化,以鼓励专业分工和创新活动。对于这两种情形,教育体系的导向似乎都是最有希望的路径:在过度多样性的社会,教育可以滋养那里急需的宽容价值观;在过度同质性的社会,教育可以提升对不同类型的有益知识的文化接受度。
3.6常见的误解
关于人际多样性对不同社会经济发展成就有何影响的研究,已吸引了经济学专业之外更广泛学术界的关注,由于方法论的差别,不出意料地招来了某些毫无根据的批评。来自文化人类学的专家提出了三类主要批评意见。以下将指出,这表明他们对相关的理论架构、统计方法、分析范围以及政策启示存在某些根本误解。
3.6.1对哥伦布时代之前美洲人口密度的低估
某些学者认为,群体多样性与总体生产率之间的驼峰状关系,是由对前殖民时代人口规模估计不准的人为错误所致。特别是,他们强调对前殖民时代美洲各社会的人口规模低估是形成这一关系的原因。
但事实上,阿什拉夫和盖勒(2013a)的历史分析已考虑了前殖民时代人口密度数据受测算误差影响的可能性,并通过多种方法使这个问题不至于动摇实证发现的有效性。
•由于人口密度是因变量,传统测算误差并不会给多样性对历史发展成就的驼峰状影响的估计造成偏差。实际上,如果没有典型的测算误差,估计得出的统计显著性甚至会更高。
•如果各大洲之间在历史人口密度测算上存在系统性差异(即美洲的历史人口密度确实被持续低估),则估计结果可能出现偏差。然而,上述研究利用各大洲固定效应开展的统计分析使得多样性对历史发展成就的影响是在各大洲内部而非各大洲之间的各个社会差异中识别的。因此对美洲人口规模的系统性低估不会对结论产生影响。
•利用公元1500年时的城市化水平而非人口密度测算历史发展成就,并不会从性质上改变人际多样性对历史发展成就的驼峰状影响。如表1第3~4列的回归结果,以及图6B所示。(*需要提示一点,公元1500年时的城市化率的数据来源同历史人口密度的数据来源无关。)
•阿什拉夫、盖勒和克莱姆普(2020a)的研究指出,人际群体多样性对若干历史时点(公元前10000年到公元1500年)人口密度的显著驼峰状影响是基于大量民族祖居地的数据(扩大样本来自Pemberton、DeGiorgio and Rosenberg,2013)。其中部分发现可参见表3第1~8列,以及图7A~图7D。
•阿什拉夫和盖勒(2013a)的主要分析是针对当地发展水平的差异,因变量为公元2000年的人均收入(而非历史人口密度)。另外,人际群体多样性对当代发展水平(即2010—2018年的平均人均收入)的驼峰状影响在回归结果中显而易见,可参见表1第5~6列,以及图6C。
•根据表1第7~8列和表3第9~10列的回归结果,以及图6D和图7E,如果采用另一种当代经济发展指标(调整后的人均夜间照明亮度),人际群体多样性的驼峰状影响依然稳健。数据来自扩展的民族样本(Pemberton、DeGiorgio and Rosenberg,2013),无论对各国或各民族祖居地均是如此。总之,对哥伦布时代之前美洲人口密度的潜在低估及其带来干扰的担忧,都是源于从根本上误解了本研究领域采用的统计方法。另外,人际群体多样性的驼峰状影响对如下几个指标均存在,进一步凸显了上述批评意见的肤浅:(1)公元1500年时的城市化率;(2)公元前10000年到公元1500年广泛区间中多个时点的民族人口密度;(3)当代的人均收入水平指标(哥伦布之前时代的潜在人口密度测算误差与此完全不相关)。
3.6.2从中性基因标记多样性到社会发展成就的映射
对本研究领域提出的另一类质疑是,中性基因标记的期望杂合度(民族内部人际多样性的代理变量)并不反映功能标记(表型标记)的多样性,因此不会影响行为与社会方面的交互作用。
HGDP-CEPH样本中的53个民族以及扩展数据库(Pemberton、DeGiorgio and Rosenberg,2013)中200多个民族的观测遗传多样性,的确都是基于中性基因标记测算得出的,所以并不直接反映功能(表型)标记的多样性。可是,有关研究的核心分析并非基于观测多样性,而是预测多样性,主要由当代民族的祖先群体的迁徙距离决定(Ashraf and Galor,2013a)。特别是如第3.3.1节所述,为克服样本限制与观测多样性带来的潜在反向因果关系问题,我们着重考察了迁徙距离对HGDP-CEPH样本中各民族观测多样性的影响,以便能够根据所有社会在前殖民时代的地理位置、相对于东非及彼此的距离,对其人际多样性水平做出预测。
关键在于,因为迁徙距离对群体内的各种表型多样性有负向影响,所以预测的人际多样性可以充当表型表达和行为表达遗传特性多样性的有效代理变量。如第3.2节所述,来自体质人类学与认知人类学的证据均显示,起源于东非的古老顺序奠基者效应影响了民族内各类形态与认知多样性在世界各地观察到的分布状况,包括与颅骨特征(Manica et al.,2007;von Cramon-Taubadel and Lycett,2008;Betti et al.,2009)、牙齿特征(Hanihara,2008)、骨盆特征(Betti et al.,2013)和产道形态特征(Betti and Manica,2018)有关的人际多样性,以及语言音素的多样性(Atkinson,2011),等等。
所以,如果认为HGDP-CEPH样本的多样性测算是基于中性基因标记,故而不能推导出人际多样性对社会经济发展成就的影响,这同样是源于他们误解了相关研究(Ashraf and Galor,2013a)的实证策略。因为迁徙距离既影响中性基因标记的多样性,又影响表型表达的形态与认知遗传特性的多样性,分析预测多样性的实证策略非常适合用来反映人际多样性对社会经济发展成就的影响。
3.6.3政策启示
某些批评者担忧,中等多样性水平使生产率最大化的结论可能导致令人不安的政策建议,例如强制推行对人口的“工程设计”,以实现人口结构中的最优多样性水平。
认为我们的主要发现会导致糟糕的政策建议,这同样是因为从根本上误读了本研究方向的核心观点及关键政策启示。人们忽略了如下主要观点:多样性对生产率的影响是通过多种机制发挥作用的,因此政策可以着眼于这些干预渠道使各个社会改变现有多样性水平实现生产率最大化的背景条件。例如第3.5节指出,鼓励多元主义的教育政策可以提升人们对不同类型的有益知识的文化接受度(这在过度同质性的社会可能较为缺乏),或者培育信任与包容的文化价值观(这在过度多样性的社会可能较为缺乏)。


















 京公网安备 11010502034662号
京公网安备 11010502034662号